9月4日,抗体药物实验室周末学术沙龙由新入职博士张利杰主讲。张博士重点介绍了其博士期间的研究工作。
MHC是一组编码主要组织相容性抗原的基因群,存在于几乎所有有颌脊椎动物的基因组中,其中包括参与动物机体特异性细胞免疫应答和体液免疫应答的经典MHC-I和MHC-II基因。尤其是MHC-II分子,在特异性抗体免疫应答中起着至关重要的作用。MHC-II分子通过识别、编辑、呈递抗原肽,形成抗原肽-MHC-II三分子复合体(pMHC-II),将抗原表位呈递给CD4+ 辅助性T细胞(Th),从而激活Th的增殖、分化,促使Th细胞分泌各种白细胞介素,白细胞介素与相应受体的结合,进一步促使B细胞分化成浆细胞并产生特异性抗体。TCR识别结合pMHC,通过CD3分子向下游传递信号。抗原肽的呈递以及TCR信号传导机制对理解细胞免疫以及新型疫苗的开发具有重要意义。
相比于MHC-I分子,MHC-II分子精细结构的研究起步较晚且难度较高。自1993年第一个MHC-II分子的结构解析至今,除了人和小鼠pMHC-II分子复合物的三维结构,没有非哺乳动物pMHC-II复合物的结构信息。鸡MHC系统最显著的现象是MHC-B区域与多种传染病(包括病毒,细菌和寄生虫)的抵抗力和敏感性相关。MHC-II分子是由α链和β链组成的异源二聚体。鸡MHC-II α链基因座即BLA位于BF-BL区域外约5 cM,无多态性。两个β链基因座BLB1和BLB2紧密连锁,具有高度多态性。鸡的MHC-II分子在基因组水平表现出了种属特征,但其呈递抗原肽的特性以及激活体液免疫应答的机制还不清楚。一条恒定的α链如何与多态性的β链配对形成功能性的异源二聚体,又是怎样呈递多样性的多肽呢。本研究通过结构生物学手段解析了鸡pMHC-II(pBL2*019:01)分子的结构,BLA链存在四个氨基酸的多出,导致pBL2*019:01具有更长的α1-β1折叠区域,同时中央界面处主链之间的氢键增加,从而稳定了PBG的底部。此外,BLB2*019:01链与鸡BLA链相互作用形成氢键的残基在已知的多态性的BLB链中高度保守。这些结果阐明了一条恒定的α链和多态性的β链配对的机制以及进化的选择性,同时也阐述了B19单倍型鸡抗原结合槽的特点,其结合多肽的特性以及其与相关分子CD4,DM之间采用不同人的作用方式。通过鸡、人、小鼠pMHC-II的结构比较,阐释了鸡MHC-II的种属特性,对于非哺乳动物MHC-II分子的研究提供了参考。
肽-MHC-I/II(pMHC-I/II)分子通过结合TCR使得CD3分子向下游传递信号而激活T细胞,并进一步促进T细胞的生物学功能。TCR及其相关蛋白复合物的结构研究在阐明T细胞机制方面具有重要的科学意义,并为疫苗开发和基于TCR的免疫疗法提供了关键指导。在过去的几十年中,对TCR的三维结构进行了广泛的研究,但仅局限于人和小鼠。本研究解析了第一个非哺乳动物鸡TCR(ch-TCRαβ)的结构。分析显示来自ch-TCRα和ch-TCRβ的两个CDR3s环之间形成带正电荷的凹槽,可能容纳朝向TCR、与MHC-I结合多肽中间部分的酸性侧链。鸡αβTCR的Cα结构域不具有人αβTCR的典型结构,由于鸡αβTCR Cα结构域的D和E片层中氨基酸数量的增加,使得DE loop环向外延伸,导致鸡αβTCR Cα结构域与CD3分子之间非常接近。另外,鸡αβTCR的Cβ结构域缺少12个氨基酸,形成了较短的FG loop环。Cα结构域DE loop环的延伸,Cβ结构域FG loop环的变短以及CD3εδ/γ堆积方向的差异证实了鸡的αβTCR和CD3εδ/γ之间具有不同与人的独特结合方式。
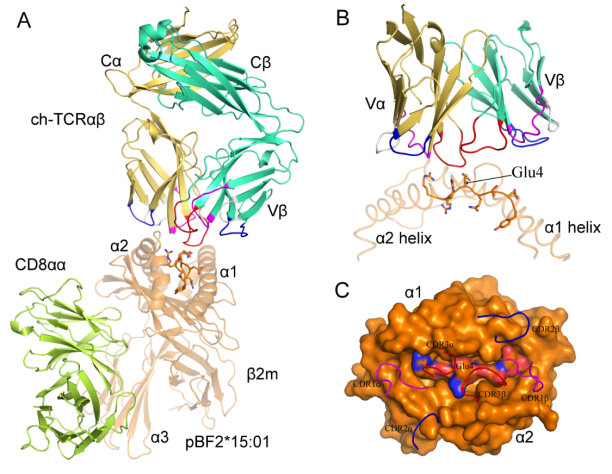
综上所述,我们解析了鸡MHC-II和αβTCR两个关键免疫分子的晶体结构。从原子水平上阐述了一条恒定的α链与多态性β链配对的结构机制,并展示了BL2*019:01呈递多肽的特性。Ch-TCRαβ结构揭示了一系列结构细节,清晰地显示了鸡αβTCR的种属特点,提出了ch-TCRαβ与MHC-I和CD3复合受体相互作用的合理描述。这些结果可推进鸡的免疫学的发展并促进疫苗学研究。